
Excerpted from Chapter 2 of Unbound
"The Technology of Spears and Digging Sticks:
Upright Posture and Bipedal
Locomotion"
NOTE: The text of this excerpt has been abbreviated. Omitted passages are indicated by ellipses ( . . . ).

The Laetoli footprints furnished indisputable evidence that early hominids were fully bipedal and have been walking on two legs for millions of years. © 2015 John Reader.
A Radical Redesign of the Mammalian Body Plan
The full upright posture and true bipedal locomotion achieved by Lucy and her kind required a wholesale transformation of the anatomy characteristic not only of all the primates but also of all mammals. This redesign included major changes to the skull, spine, pelvis, legs, feet, and other musculoskeletal structures. For the first time, the weight of the entire upper body had to be supported by the hind legs alone, an arrangement that had never before existed among any of the placental mammals. What survival advantage was powerful enough to completely reorganize the way the primate body stood, walked, and ran?
. . . The place where the skull is attached to
the spinal column, called the foramen magnum, had to move from the
back of the skull, where it is located in all other animals, to the bottom
of the skull, where it is located in humans. . . . [I]n a
two-legged mammal, the spine and pelvis must support the weight
of the entire upper body, including the head and neck, forearms and
shoulders, chest, and abdomen. . . .To accomplish this feat, the spine had to be greatly
strengthened and bent into an S curve, to bring the center of gravity of
the entire body directly over the pelvis.
In order to support the weight of the entire upper body, the pelvis also had to undergo a major restructuring. . . . the pelvic bones had to become shorter, thicker, and fused together into a single rigid, doughnut-shaped structure. At the same time, the bones of the hind legs became long and straight, with the ability to lock at the knee joints. When other primates stand on their hind legs, their knees are bent. The evolution of locking knee joints enabled the bipedal hominids to stand fully upright for long periods of time without putting constant strain on their leg muscles.

The Bonobo, our closest primate relative, can stand and walk upright when carrying things with its forearms, but its bent knees reveals that its legs are designed for quadrupedal locomotion. © Frans Lanting/www.lanting.com.
Finally, the grasping feet and opposable big toes typical of all other primates had to be completely transformed. . . . If you look at the thumb on the rear foot of a chimp or gorilla, you will see that it is located on the side of the foot and is opposable to the other toes, similar to the opposable thumb in the human hand. This is necessary for grasping with the hind legs while climbing about in tree branches. . . . So this grasping thumb migrated from its original position opposite the other four toes to a new position in which it is lined up with the other toes, allowing all five toes to face in the same direction. At this point, the ape’s thumb had become the human’s big toe.
At the same time, the palm of the foot lengthened, eventually developing a distinct arch, and the “fingers” on the hind feet became shorter, eventually turning into the four stubby little toes of our own species that are no longer capable of grasping much of anything but are perfectly capable of supporting their new function of walking and running for long distances over the open ground.
When an animal evolves a radical head-to-toe redesign of the basic body plan that served its ancestors well for tens of millions of years, especially in a short space of geological time, we can assume that powerful evolutionary forces are at work. But scientists have struggled to reach consensus about what these evolutionary forces were. In fact, a number of different theories have been proposed to explain precisely why our ancestors embarked upon the evolution of their unique and unprecedented form of upright posture and bipedal locomotion.
. . .
The early hominids lived during a period in African prehistory when the forests were shrinking and the grasslands were expanding. For a long time, it was believed that bipedal locomotion evolved primarily as an adaptation to the drier savannah environment— characterized by grasslands dotted with scattered trees—that was spreading throughout equatorial Africa. It was assumed that walking and running on two legs had enabled Lucy’s ancestors to leave their original forest habitat, see more clearly over the tops of the grasses, and cover longer and longer distances over the open ground. This is known as the “savannah-based hypothesis.” There are, however, some major problems with this theory.
First, all other grassland species . . . are strictly quadrupedal. (In fact, all land mammals except hominids are quadrupedal.) Second, of all the other primates who also made the transition to a terrestrial way of life (these include baboons, mandrills, vervet monkeys, and patas monkeys), none of them adopted upright posture. . . .
Third, the earliest anatomical evidence of bipedal locomotion comes from the fossil remains of an ape-like creature called Ardipithecus, who first appeared roughly five million years ago—at least two million years before Lucy was born. In a major setback for the savannah-based hypothesis, it has been firmly established that Ardipithecus lived and died in a forest environment and never inhabited the grasslands at all.
. . .
Due to the problems of the savannah-based theory, a variety of alternative and often competing theories have been proposed in recent years to explain the evolution of upright posture and bipedal locomotion. . . . But none of these mechanisms seems to involve the kind of life-or-death survival advantages that would have been necessary for the radical restructuring of primate anatomy that bipedalism required.
The “provisioning hypothesis” proposes that hunting males needed free hands in order to carry their prey back to the home base for the females and offspring. However, this does not account for the fact that other predatory mammals bring meat back for their offspring by simply carrying it in their mouths. While this behavior might have required an evolutionary adjustment (since primates rarely use their mouths for carrying things), it would hardly have required a complete restructuring of primate anatomy.
The “thermal loading hypothesis” proposes that by standing upright, the early hominids exposed a smaller area of their bodies to the fierce tropical sun and were thus able to remain cooler while exerting themselves out in the open. But in addition to the fact that this does not explain why no other tropical animals have ever adopted a similar strategy, we should recall that bipedalism first began with Ardipithecus, who lived largely in the shade of the tropical forests.
The “warning display hypothesis” proposes that individuals (especially males) that could threaten others by frequently standing erect were more dominant and thus sexually outcompeted individuals who stood erect less frequently. However, this hypothesis fails to account for the fact that gorillas, bears, and other species that also threaten their rivals by standing erect have never adopted bipedalism.
The “postural feeding hypothesis” proposes that bipedalism enabled prehistoric apes to stand upright while gathering low-hanging fruit with both hands, rather than hanging on to a branch with one hand and gathering fruit with the other. However, it seems unlikely that this would confer a sufficiently powerful survival advantage to produce the massive anatomical changes that bipedalism entailed. Furthermore, the essence of bipedalism is not merely standing upright but walking and running on two legs. Apes, monkeys, bears, beavers, prairie dogs, meerkats, and many other mammals can easily stand upright, but only hominids are specially adapted for bipedal locomotion.
The primary weakness of these theories is that none of them explains why the large canine teeth of our primate ancestors disappeared at the same time as bipedal locomotion evolved. Instead, the loss of large canines among the hominids is typically explained by the “diminished male rivalry hypothesis,” which proposes that large canines disappeared in order to prevent adult male hominids from engaging in deadly combat, because this might have interfered with their ability to cooperate and become more effective hunters.
However, lions, wolves, hyenas, chimpanzees, and many other species that engage in cooperative hunting do not have diminished biological weapons. Besides, male humans are entirely capable of both close cooperation and deadly conflict with each other, and the absence of large canines has not prevented male humans from engaging in deadly conflict throughout all of human history. Instead of using their teeth to kill each other, they just use their lethal weapons.
So, the question remains. How could any of the early hominids have survived for millions of years in an environment full of large and dangerous predators without any effective way of defending themselves?
The Disappearing Canine Teeth
The canine teeth of Ardipithecus ramidus—the earliest species to display
skeletal evidence of bipedal locomotion—had already become significantly
diminished compared to the canines of its ancestors. By the
time of Lucy and the early hominids, the large, deadly, weapons-grade
canine teeth possessed by all other primate species had completely disappeared,
never to be seen again among the hominids.
In fact, one of the members of the team that investigated the Ardipithecus ramidus site observed that male canine reduction was well under way by six million years ago. The loss of weapons-grade canine teeth began millions of years before Lucy appeared, and by the time of the early hominids these large canine teeth—the only biological weapon the primates ever possessed during their entire fifty-million-year history— had completely disappeared.
No monkey, ape, or hominid has claws, horns, antlers, hooves, or tusks. But they all have formidable canine teeth that can inflict painful and even lethal damage to an adversary. Yet the Australopithecus fossils have only small, dull, flat-wearing canine teeth that could not possibly have competed with the dangerous biological weapons of their natural enemies.
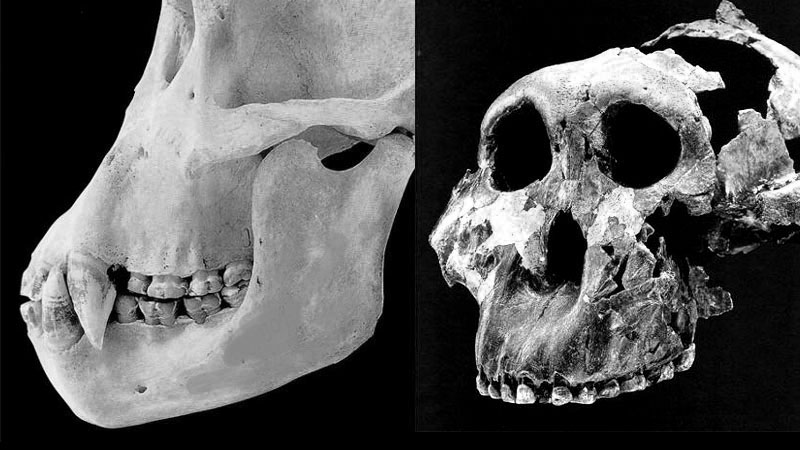
Compare the the weapons-grade canines of our closest relative, the common chimpanzee (LEFT) with the blunt, stubby canine teeth of the early hominid Paranthropus boiseii (RIGHT). © 1985, 1996 David L Brill
Furthermore, apes and monkeys all have superior tree-climbing abilities that enable them to climb high into the treetops, far beyond the reach of their natural enemies like the big cats. Yet the long, straight legs and shrunken toes of the australopithecines and the other hominid species who followed them would have seriously reduced their ability to climb out of danger, making them that much more vulnerable to their natural enemies.
A species’ natural defenses disappear only when they are no longer needed for survival. Although primates can certainly throw stones, a stone heavy enough to cause real damage to a large predator would be too heavy to throw any distance with any great accuracy. . . . Heavy clubs made from large branches can crush the skull of an adversary, but only at close range. If you get close enough to a predator to hit it over the head with a club, it will probably be close enough to maul you with its claws, and—unless you have dispatched it with a single blow—it will already have its fangs at your throat. How did the early hominids compete so successfully against “nature red in tooth and claw” with neither the teeth nor the claws to fight back, when all they had to fight with was their “bare hands”?
The most likely answer is that the hands of the early hominids were not bare. They must have been carrying weapons. And the weapons they fashioned to attack both predators and prey must have been so superior to the formidable canine teeth of their primate ancestors that having effective dental weaponry was no longer necessary. As we saw in chapter 1, the use of technology is not limited to hominids. Certain wild chimpanzees have even been observed making primitive spears that they use for killing their prey. It is therefore most likely that when prehistoric apes began to make spears, they unleashed the evolutionary forces that eventually produced an ape with full upright posture and bipedal locomotion.
. . .
The spear would have completely changed the game of hunting and defense, because long spears can be used with lethal force while hunters remain safely out of reach of the biological weapons of their prey. If a group of hunters can impale an animal with spears that are several feet long, it can be killed without ever coming close enough to use its biological weapons on its attackers.
. . .
The ability to carry and wield a spear long enough to attack and kill another animal from beyond the range of its biological weaponry would have had powerful survival value. Once the ancestors of the hominids had adopted the technology of the spear, those individuals who could stand firmly on their hind limbs—while jabbing and thrusting a spear with their forelimbs—would have had a clear advantage over their competitors. The longer these individuals could remain standing, the farther they could walk and run on two legs, and the larger and heavier weapons they could carry with them, the more effectively they could defend their mates and offspring against attacks from would-be predators—and the more meat they could bring back to share with the other members of the group. For all of these reasons, the survival value of fabricated spears would have been sufficient to bring about the major anatomical changes required for a four-footed animal to evolve into a two-footed animal.
. . .
The loss of dental weaponry among all prehistoric bipeds proved to be complete. None of the early hominids ever regained the large and dangerous canines of their prehistoric ancestors. When the technology of making and using spears was adopted by an ancient population of prehistoric apes, it became a strategy for hunting and defense that effectively trumped the biological weaponry of both the predators and prey living in those prehistoric environments.
Once the hind legs were capable of assuming full responsibility for locomotion, the forelegs were free to serve other purposes. They could be used to make tools and weapons, and they could also be used to carry these tools and weapons from place to place. The bipedal hominids were able to use their free hands and arms to transport relatively heavy burdens—such as the spoils of the hunt or a trove of ripe fruit— from one place to another, making it possible to bring food that was acquired in distant locations back to a communal home base, as the provisioning hypothesis suggested. And the same basic technology—in the form of a digging stick—would have also been used to gather foods that are buried in the earth. In hunting and gathering societies, women routinely used such sticks to dig up roots, bulbs, tubers, termite nests, and the burrows of small animals, as well as to knock down nuts and fruits from the ends of tree branches that were too slender to climb.
In the final analysis, it appears that the technology of spears and digging sticks not only stimulated the evolution of bipedalism but also was ultimately responsible for the evolution of humanity itself. However outlandish this claim may seem, the human body’s unique, much loved, and much celebrated physical form most likely owes its origin to a group of ancient apes—long since lost in the mists of prehistory— that first mastered the extraordinary offensive, defensive, and food-gathering possibilities of the long, sharp stick.